Bacteria exposed to antiviral drugs develop antibiotic cross ... - Nature.com
Abstract
Antiviral drugs are used globally as treatment and prophylaxis for long-term and acute viral infections. Even though antivirals also have been shown to have off-target effects on bacterial growth, the potential contributions of antivirals to antimicrobial resistance remains unknown. Herein we explored the ability of different classes of antiviral drugs to induce antimicrobial resistance. Our results establish the previously unrecognized capacity of antivirals to broadly alter the phenotypic antimicrobial resistance profiles of both gram-negative and gram-positive bacteria Escherichia coli and Bacillus cereus. Bacteria exposed to antivirals including zidovudine, dolutegravir and raltegravir developed cross-resistance to commonly used antibiotics including trimethoprim, tetracycline, clarithromycin, erythromycin, and amoxicillin. Whole genome sequencing of antiviral-resistant E. coli isolates revealed numerous unique single base pair mutations, as well as multi-base pair insertions and deletions, in genes with known and suspected roles in antimicrobial resistance including those coding for multidrug efflux pumps, carbohydrate transport, and cellular metabolism. The observed phenotypic changes coupled with genotypic results indicate that bacteria exposed to antiviral drugs with antibacterial properties in vitro can develop multiple resistance mutations that confer cross-resistance to antibiotics. Our findings underscore the potential contribution of wide scale usage of antiviral drugs to the development and spread of antimicrobial resistance in humans and the environment.
Introduction
Antiviral drugs are used worldwide to treat viral diseases that affect millions of human lives. Antivirals attenuate viral infection by inhibiting a virus's ability to replicate, often by targeting the proteins or enzymes used by a virus to infect, multiply, or release new viral particles from a host1. Ninety antivirals are currently (as of 2017) approved by the Food and Drug Administration in the United States, including single-compound and combination antivirals for influenza virus, hepatitis B and C, herpes simplex virus 1 and 2, and human immunodeficiency virus (HIV)2. Worldwide, an estimated 38 million people are infected with HIV3. Three hundred twenty-five million people are living with hepatitis B and/or C4, and over 3.7 billion cases of herpes simplex virus type 1 exist in individuals under the age of 50 alone5. An estimated three to five million severe cases of influenza virus infection occur annually worldwide6. As the number of cases of viral diseases increases every year, the use of antiviral drugs is expected to rise as well7. Some viral infections such as seasonal influenza may be treated with only short-term antiviral therapy; others such as HIV/AIDS can require long-term to lifetime antiviral treatment. In addition, the on-going SARS-CoV-2 pandemic adds to the list of global viral diseases treated with antiviral drugs8.
Given the extensive use of antiviral drugs worldwide, the unforeseen consequences of antiviral drugs must also be considered. Antiviral drugs are intended to target viral replication specifically; however, antivirals can have off-target effects including the inhibition of bacterial growth9,10,11,12. The antibacterial activity of several antiviral drugs has led to increased interest over the last decade in repurposing nucleoside analog antivirals, in particular, to treat multidrug resistant bacterial infections13,14,15. However, the antibacterial activity of many antiviral drugs also raises the question of whether antivirals can contribute to antimicrobial resistance, which encompasses the presence, development, dissemination, and treatment of antimicrobial resistant infections. Globally, antimicrobial resistance is one of the top ten threats to human health16. In the United States alone, there are over 2.8 million antibiotic-resistant infections every year and over 35,000 deaths due to antibiotic resistance17. Most discussion on antimicrobial resistance control centers on the judicious use of prescription antibiotics and hygiene18,19. However, an understanding of the full spectrum of contributors to antimicrobial resistance remains incomplete. Non-antibiotic pharmaceuticals such as antivirals may also contribute to the development of antimicrobial resistance on a global scale, but research is needed to gain full understanding of how drugs with antibacterial properties can lead to mutations in bacteria and cause cross-resistance to antibiotics.
While the antibacterial properties of many antiviral drugs have been demonstrated, it is largely unknown whether bacteria can become resistant to these antivirals and whether resistance mutations to antibacterial antivirals can confer cross-resistance to other antibacterial agents including antibiotic drugs. Non-pharmaceutical compounds with antimicrobial effects, such as some pesticides and herbicides, have been shown to contribute to antimicrobial resistance20,21,22. Other non-antibiotic drugs such as antidepressants and antiepileptic drugs have been shown to ultimately influence resistance by increasing rates of horizontal gene transfer, affecting the same or similar targets as antibiotic drugs, or resulting in changes to gene expression impacting resistance to antibiotic drugs23,24,25. Therefore, examining both the phenotypic and genotypic impacts of non-antibiotic drug exposure such as antivirals on bacteria can lead to better understanding of cross-resistance and the mechanisms leading to resistance. Beyond commonly known genetic mutations that cause antibiotic resistance, novel mutations due to non-antibiotic drug exposure may be responsible for conferring antibiotic cross-resistance26. By conducting whole genome sequencing, novel resistance mutations can be identified to help explain resistance phenotypes27,28.
In this study, we investigated the antibacterial efficacy of antivirals and the potential for cross-resistance to other antivirals and antibiotic drugs. Using a culture-based approach with the model bacteria E. coli and B. cereus, we demonstrate that antiviral drugs have the potential to contribute to antibiotic resistance in gram-negative and gram-positive bacteria. While other studies have previously demonstrated the antibacterial properties of some antiviral drugs, to our knowledge, no other work has yet reported the extent to which numerous antivirals of different classes may lead to antibiotic cross-resistance. The results of this study suggest that antiviral drugs with antibacterial properties can impact the antimicrobial resistance phenotype of bacteria; further investigation may help elucidate the specific mechanisms by which these drugs and potentially others may act on bacteria and contribute to antibiotic cross-resistance.
Results
Antibacterial effects of antivirals on 24-h growth of E. coli and B. cereus
Fourteen antiviral drugs from four different antiviral classes (antiherpetic, nucleoside reverse transcriptase inhibitor (NRTI), integrase inhibitor (II), non-nucleoside reverse transcriptase inhibitor (NNRTI)) with a range of different targets/ modes of action (Supplementary Data 1) were screened for antibacterial activity against E. coli and B. cereus.
Demonstrating consistent results with previous work10, eight of the fourteen antivirals tested inhibited the growth of E. coli, while only three of the fourteen displayed antibacterial activity against B. cereus (Fig. 1). The antivirals abacavir, darunavir, nevirapine, tenofovir, favipiravir, and emtricitabine had no significant impact on the growth of either E. coli or B. cereus up to 100 μg/mL over 24 h (Supplementary Fig. 1, Supplementary Fig. 2). Dolutegravir, efavirenz, and raltegravir significantly reduced the growth of both E. coli and B. cereus, while the nucleoside analogs acyclovir, didanosine, lamivudine, stavudine, and zidovudine significantly inhibited E. coli but not B. cereus (Fig. 1). Among the antivirals that displayed antibacterial activity, growth inhibition was typically observed at concentrations ≥ 50 μg/mL; however, zidovudine significantly inhibited E. coli growth at 0.1 μg/mL (Fig. 1). Growth inhibition, as seen by a significant reduction in optical density (OD), was observed within 4 h of exposure for several of the antivirals with antibacterial activity against E. coli, and all antivirals with antibacterial activity inhibited growth within 8 h(Fig. 1). In some cases, the significance of antimicrobial effect was maintained over time, while in other cases it changed over time, either increasing or decreasing (Fig. 1). We were conservative with what we considered a significant inhibition of growth, and we required both significant (p < 0.005) and substantial (>10%) reduction in growth of the antiviral-treated bacteria compared to untreated controls (Supplementary Data 3, Supplementary Fig. 2). Differences were determined significant if the p-value of the t-test met the threshold (p < 0.005). All significant differences were further differentiated according to their degree of significance (from most to least significant: p < 0.00005, p < 0.0005, p < 0.005) for comparative purposes. Results that were not significant were interpreted as no significant difference in growth between the drug-treated condition and the untreated condition.

a Antibacterial effects of antivirals after 4–24 h of exposure at different antiviral concentrations ranging from 0.1–100 μg/mL. Nucleoside analogs acyclovir, didanosine, lamivudine, stavudine and zidovudine impact the growth of E. coli only. Both E. coli and B. cereus are significantly inhibited (with at least 10% reduction in growth compared to control) by non-nucleoside reverse transcriptase inhibitor efavirenz and the integrase inhibitors dolutegravir and raltegravir at concentrations from 10–100 μg/mL beginning at 4 h of exposure time for E. coli and 8 h for B. cereus. b Example of growth curves for E. coli treated with 0.1–100 μg/mL acyclovir over 24 h. The most significant difference in growth, as observed in the growth curve and noted in the heat map (a) is only from 8–24 h with 100 μg/mL acyclovir. c Example of growth curves for E. coli treated with 0.1–100 μg/mL zidovudine over 24 h. Significant differences between untreated and zidovudine-treated E. coli are observable at all concentrations tested and across all time points from 4–24 h.
Antiviral resistance and cross-resistance after repeated antiviral exposure
After demonstrating that eight of the tested antivirals had significant antibacterial effects on the growth of E. coli and three impacted the growth of B. cereus, we explored the potential for bacteria to become resistant to the antibacterial effects of these antivirals upon repeated exposure to the investigated drug. Antiviral-resistant strains that were isolated included zidovudine-resistant, lamivudine-resistant, stavudine-resistant, and didanosine-resistant E. coli. Dolutegravir-resistant and raltegravir-resistant strains were isolated for both E. coli and B. cereus. Resistant strains achieved similar maximum growth and growth kinetics compared to wild type, with the exception of the E. coli zidovudine-resistant strain which achieved 32% lower maximal growth compared to wild type (Supplementary Data 4; Supplementary Fig. 3). During the incubation period of 4–12 h, while bacteria were in log phase and OD was rapidly increasing, wild type B. cereus showed an average increase in OD of 0.162 (absorbance, 600 nm) per hour, and the antiviral-resistant strains showed an increase of 0.154 per hour. Wild-type E. coli showed an OD increase of 0.162 per hour, and antiviral-resistant strains (excluding zidovudine-resistant E. coli) showed an average increase in OD of 0.199 per hour (Supplementary Data 4). Resistance to antivirals was observed at sub-inhibitory concentrations, typically at <100 μg/mL (Supplementary Data 5; Supplementary Fig. 4). Resistance was not observed in efavirenz treatments for both E. coli and B. cereus; resistance was observed in E. coli acyclovir treatments, but a resistant strain was not isolated (Supplementary Data 5).
Resistance to one antiviral drug conferred cross-resistance to other antiviral drugs for all six of the E. coli and both B. cereus resistant strains developed (Fig. 2, Supplementary Data 6). Although each strain displayed a unique pattern of cross-resistance to other antivirals, cross-resistance profiles clustered in some cases by the antiviral class from which each strain was isolated (Supplementary Fig. 5). For example, E. coli mutants isolated from the NRTI treatments lamivudine and stavudine were cross-resistant to the NRTIs lamivudine, stavudine and didanosine but susceptible to zidovudine (NRTI), dolutegravir (II), and efavirenz (NNRTI). These strains clustered together and were also similar to the NRTI didanosine-resistant strain (whose profile was more closely related to wild type E. coli rather than the lamivudine- and stavudine-resistant strains). Likewise, strains resistant to integrase inhibitors dolutegravir and raltegravir displayed similar cross-resistance profiles and clustered together. These strains were characterized by broader cross-resistance than the NRTI-resistant strains and were resistant to all tested antivirals except zidovudine. Despite being an NRTI like lamivudine, stavudine and didanosine, the zidovudine-resistant E. coli strain exhibited a uniquely broad cross-resistance profile that was more similar to the resistance profiles of the integrase inhibitor-resistant strains but stood out uniquely. Zidovudine-resistant E. coli was the only nucleoside-analog-resistant strain to show cross-resistance to NNRTI efavirenz compared to wild type (40% growth compared to 18% growth for wild type E. coli) as well as zidovudine itself.

a Compared to wild type E. coli, the antiviral-resistant strains show cross-resistance to multiple antivirals, and unique resistance profiles are observed for all E. coli mutant strains resistant to NRTIs (didanosine-resistant, Didanosine-R; lamivudine-resistant, Lamivudine-R; stavudine-resistant, Stavudine-R; zidovudine-resistant, Zidovudine-R) and integrase inhibitors (dolutegravir-resistant, Dolutegravir-R; raltegravir-resistant, Raltegravir-R). b Dolutegravir-R and Raltegravir-R B. cereus strains are resistant to efavirenz and dolutegravir compared to wild type B. cereus.
Isolated E. coli and B. cereus strains resistant to integrase inhibitors dolutegravir and raltegravir demonstrated similarities in their cross-resistance to efavirenz, dolutegravir and raltegravir (Fig. 2a, b). Raltegravir-resistant B. cereus were resistant to efavirenz compared to wild type B. cereus (93% growth versus 9% growth). Interestingly, the raltegravir-resistant B. cereus mutant strain showed greater resistance to dolutegravir compared to the "dolutegravir-resistant strain" that was developed in the presence of dolutegravir and isolated for cross-resistance testing (Fig. 2b). Antiviral-resistant strains were, overall, shown to have stable resistance phenotypes, as demonstrated over 15 passages. For example, dolutegravir-resistant B. cereus maintained resistance to raltegravir, dolutegravir, and efavirenz throughout passaging in the absence of dolutegravir; zidovudine-resistant E. coli maintained resistance to zidovudine throughout 15 passages in absence of zidovudine (Supplementary Data 7).
Antibiotic cross-resistance in antiviral-resistant E. coli and B. cereus
We further explored the extent of acquired resistance in E. coli and B. cereus by challenging the isolated antiviral-resistant strains with ten commonly prescribed broad-spectrum antibiotics (Supplementary Data 8). The final concentrations of antibiotics used to challenge wild type and mutant E. coli and B. cereus were selected after first challenging each individual strain and then choosing a concentration that captured the range of response to drug—i.e., changes in growth– across wild type and antiviral-resistant mutant strains for comparative analysis. Similar to their cross-resistances with other antivirals, antiviral-resistant E. coli and B. cereus strains each exhibited unique resistance/susceptibility patterns to antibiotics (Fig. 3, Fig. 4, see Supplementary Data 9 for determinations of significance). Some mutant strains demonstrated strong resistance to antibiotics, and some strains were more susceptible to antibiotics compared to wild type control. E. coli and B. cereus mutants resistant to the same antiviral exhibited different antibiotic cross-resistance. For example, dolutegravir-resistant E. coli showed some resistance to tetracycline and oxytetracycline (Fig. 3) while dolutegravir-resistant B. cereus were more susceptible to tetracycline and oxytetracycline compared to wild type (Fig. 4).
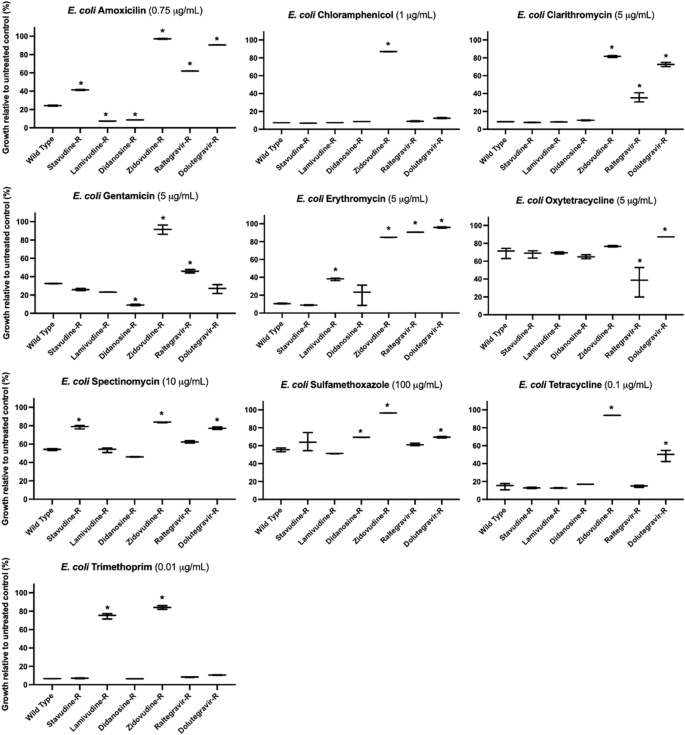
However, some antiviral-resistant E. coli mutants are more susceptible to some antibiotics compared to wild type E. coli, including increased susceptibility of didanosine-resistant (Didanosine-R) E. coli to amoxicillin and gentamicin. Asterisks (*) indicate significant difference compared to wild type (p < 0.05 and at least 10% substantial difference in growth between treated wild type and resistant strain); bars represent maximum, minimum and mean values for n = 3 absorbance measurements.
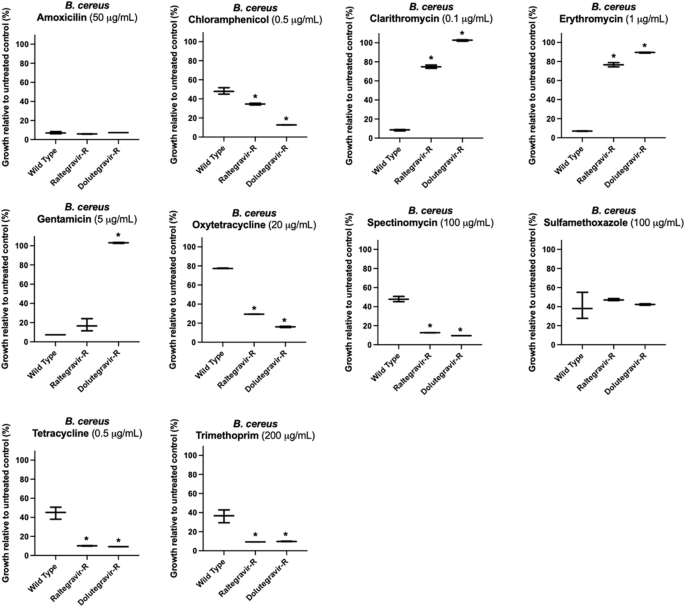
However, B. cereus antiviral-resistant mutants raltegravir-resistant (Raltegravir-R) and dolutegravir-resistant (Dolutegravir-R) were often equally or more susceptible to antibiotics compared to wild type B. cereus, as illustrated by their response to oxytetracycline or spectinomycin. Asterisks (*) indicate significant difference compared to wild type (p < 0.05 and at least 10% substantial difference in growth between treated wild type and resistant strain); bars represent maximum, minimum and mean values for n = 3 absorbance measurements.
Most antiviral-resistant E. coli mutant strains exhibited several strong antibiotic cross-resistances. Raltegravir-resistant, dolutegravir-resistant, and zidovudine-resistant E. coli showed resistance to clarithromycin and erythromycin (macrolide antibiotics) (Fig. 3). Zidovudine-resistant and lamivudine-resistant E. coli were resistant to trimethoprim. Dolutegravir-resistant and zidovudine-resistant E. coli strains showed cross-resistance to tetracycline. Compared to wild-type E. coli, dolutegravir-resistant E. coli also showed decreased susceptibility to amoxicillin, spectinomycin, and sulfamethoxazole (24 vs 90%, 54 vs 77%, 56 vs 70%, wild type vs dolutegravir-resistant growth, respectively) in addition to the macrolide and tetracycline antibiotics (Fig. 3, Supplementary Data 10). Some antiviral-resistant E. coli strains were more susceptible to antibiotics compared to wild type E. coli. Greater susceptibility was observed for lamivudine-resistant and didanosine-resistant E. coli to amoxicillin and raltegravir-resistant E. coli to oxytetracycline. When treated with gentamicin (aminoglycoside), both raltegravir-resistant and zidovudine-resistant E. coli were more resistant compared to wild type E. coli (46% vs 32% growth for raltegravir-resistant, and 92% vs 32% for zidovudine-resistant E. coli).
B. cereus raltegravir-resistant and dolutegravir-resistant strains were either more susceptible or comparably susceptible to seven out of ten antibiotics tested compared to the wild type B. cereus. Both dolutegravir-resistant and raltegravir-resistant B. cereus were equally susceptible to amoxicillin and sulfamethoxazole compared to wild type (Fig. 4). The strains were more susceptible to spectinomycin, tetracycline, trimethoprim, oxytetracycline and chloramphenicol compared to wild type B. cereus (Fig. 4). However, for the antibiotics gentamicin, erythromycin and clarithromycin, the antiviral-resistant strains showed greater resistance compared to wild type B. cereus: only dolutegravir-resistant B. cereus was resistant to gentamicin; similar to dolutegravir-resistant and raltegravir-resistant E. coli, dolutegravir-resistant and raltegravir-resistant B. cereus both were resistant to clarithromycin and erythromycin. When treated with 1 μg/mL erythromycin, wild type B. cereus exhibited on average only 7% growth relative to untreated wild type, while dolutegravir-resistant B. cereus exhibited 90% growth relative to untreated dolutegravir-resistant. Raltegravir-resistant B. cereus exhibited 76% growth relative to untreated raltegravir-resistant in the presence of erythromycin. When exposed to 0.1 μg/ml clarithromycin, wild type B. cereus exhibited 9% growth while dolutegravir-resistant and raltegravir-resistant strains exhibited 103% growth and 75% growth, respectively.
Comparative phenotypic analysis of multidrug-resistant E. coli
Following our results demonstrating that antiviral-resistant E. coli can exhibit broad cross-resistance to antibiotic drugs, we wanted to contextualize the clinical relevance of the survival and growth of antiviral-resistant E. coli. To validate the antibiotic cross-resistance phenotypes of the antiviral-resistant strains and the in vitro method of evaluation for resistance, we challenged a well-characterized multidrug-resistant E. coli strain (ATCC BAA-2471, clinical respiratory sample) with the same antibiotics and concentrations used to challenge antiviral-resistant E. coli strains. E. coli BAA-2471 exhibits resistance to many classes of antibiotics including penicillins, cephalosporins, carbapenems, quinolones, aminoglycosides, and others29. The strain is therefore resistant to antibiotics we tested including amoxicillin, gentamicin, tetracycline, trimethoprim and sulfamethoxazole. We confirmed the resistance of BAA-2471 to these antibiotics using our methods, and we also compared the strain's resistance or susceptibility to antiviral drugs (Fig. 5, Supplementary Data 11). Zidovudine-resistant E. coli—the antiviral-resistant E. coli strain with the broadest resistance profile—and untreated BAA-2471 exhibited comparable growth kinetics. Resistance was indeed observed for all antibiotics to which BAA-2471 was purportedly resistant, and the growth of BAA-2471 and zidovudine-resistant E. coli in the presence of antibiotic treatment was comparable (Fig. 5, Supplementary Data 15). While both BAA-2471 and zidovudine-resistant E. coli demonstrated growth in the presence of 3 μg/ml gentamicin, we found growth of BAA-2471 inhibited at 16 h' incubation with 5 μg/ml gentamicin in our assays whereas zidovudine-resistant E. coli were not susceptible (Fig. 3, Supplementary Data 11). In the presence of 50 μg/ml zidovudine, after 16 h' incubation, zidovudine-resistant E. coli exhibited 120% growth compared to untreated control whereas BAA-2471 exhibited only 48% growth (Fig. 5, Supplementary Data 15). These results both validated our approach to assessing the phenotypic resistance profiles of resistant strains and allowed comparative analysis of bacterial growth in the presence and absence of antiviral drugs and antibiotics using a well-characterized resistant clinical isolate.

As expected, multidrug-resistant clinical E. coli isolate BAA-2471 is resistant to amoxicillin and tetracycline at the concentrations tested, while wild type E. coli were susceptible at these concentrations (wild type response shown in Fig. 3). The growth curves of zidovudine-resistant E. coli (AZT-R) demonstrate the same resistance as BAA-2471 to the antibiotics as well as the antiviral dolutegravir. However, BAA-2471 is susceptible to zidovudine. For each time point, n = 3 technical replicates; mean absorbance is plotted for each time point, and absence of observable error bars at any data point is indicative of close alignment of triplicate measurements.
Further, the results suggest that zidovudine-resistant E. coli exhibit resistance mutations that are unique compared to the clinical isolate BAA-2471 yet grant comparable fitness in the absence of drug and comparable survival in the presence of antibiotic concentrations that are inhibitory for wild type.
Comparative genome analysis of antiviral-resistant E. coli
After observing that antiviral-resistant isolates of E. coli and B. cereus have phenotypically unique changes in resistance to antibiotics compared to wild type, we conducted whole genome sequencing of antiviral-resistant E. coli strains with the goal of identifying genetic changes that may explain resistance to either antiviral drugs or antibiotics. Comparative genomics of B. cereus isolates could not be conducted due to contamination during reference strain library preparation and small number of mutant strains. Overall, there were as few as two and as many as 23 unique genetic changes in the antiviral-resistant E. coli mutant genomes as compared with the reference strain that could not be attributed to either a change in the reference during sequencing (differences found in all mutant genomes) or to a sequencing error (e.g., insertion of A within A homopolymer sequence) (Supplementary Data 13). Mutations observed in three of the mutant strains (raltegravir-resistant E. coli, [CP117043]; dolutegravir-resistant E. coli, [CP117044]; didanosine-resistant E. coli, [CP115968]; Fig. 6, Fig. 7) had genetic changes in genes with established roles in antibiotic resistance. Raltegravir-resistant and dolutegravir-resistant E. coli isolates—both integrase inhibitor resistant strains– have the same base change (A→G) leading to a non-synonymous amino acid change (Y→C) in the cAMP-activated global transcriptional regulator crp (Fig. 7), associated with resistance to macrolides, potentially explaining part of their acquired antibiotic resistance phenotype of these two antivirals from the same antiviral class. A mutation in the multidrug efflux pump, efflux resistance-nodulation-division (RND) transporter permease acrB, was observed in the dolutegravir-resistant E. coli isolate. In this case, the observed mutation led to a 1.8% increase in the length of the protein (Fig. 7) and may explain the observed resistance to tetracycline30,31,32,33. In didanosine-resistant E. coli, a deletion was observed in ABC transporter permease yhdY (single base pair deletion in codon 232), suggesting potentially altered function to this protein family involved in the transport of a variety of substituents including not only carbohydrates, proteins, and lipids but also pharmaceutical drugs34 (Fig. 6). The early stop codon introduced by the single base pair deletion led to a 35.2% reduction in protein length. Additionally, two mutants exposed to nucleoside analogs (lamivudine-resistant E, coli, [CP115179], cytidine analog; zidovudine-resistant E, coli, [CP117469], thymidine analog) had different genetic changes within thymidine kinase gene (tdk), suggesting a role in nucleoside analog antiviral drug resistance (Supplementary Data 13).

Blue highlighting indicates known role in antimicrobial resistance; green highlighting indicates recently suggested role in resistance. +Accession number listed below each genome. *"Mutation" column lists first in parentheses the codon number in the wild type reference gene that is mutated compared to the antiviral-resistant strain, followed by the mutation described as the single letter amino acid abbreviation and sequence of the wild type and the single letter amino acid abbreviation and altered sequence in the antiviral-resistant mutant gene.

Blue highlighting indicates known role in antimicrobial resistance; green highlighting indicates recently suggested role in resistance. +Accession number listed below each genome. *"Mutation" column lists first in parentheses the codon number in the wild type reference gene that is mutated compared to the antiviral-resistant strain, followed by the mutation described as the single letter amino acid abbreviation and sequence of the wild type and the single letter amino acid abbreviation and altered sequence in the antiviral-resistant mutant gene. #The protein increased in length by 1.8% due to the observed mutation.
Other mutations causing substantial changes in protein sequences observed in antiviral-resistant E. coli were located in genes implicated in antibiotic resistance in recent studies. For example, mutations were observed in the coding region for the phosphotransferase system (PTS) N-acetylgalactosamine-specific transporter subunit IIB (agaV) in dolutegravir-resistant E. coli. This system is involved in carbohydrate transport, yet mutations in PTS have also been associated with pan-tolerance to antimicrobials26 (Fig. 7). The mutation in agaV led to 47.6% reduction in the length of the protein. Also, in dolutegravir-resistant E. coli, a five-base pair insertion was observed in the sensory domain of two-component sensor kinase family protein, which consists of a histidine kinase and cognate response regulator. Various mutations in this family of proteins have been linked to antimicrobial resistance35. Lamivudine-resistant E. coli exhibited a single base pair deletion in the region coding for hexose-6-phosphate:phosphate antiporter (uhpT), leading to 73.1% truncation of the protein (Fig. 6). This antiporter is involved in the exchange of hexose-6-phosphate (which could include the six-carbon sugars glucose-6-phosphate, fructose-6-phosphate and mannose-6-phosphate) and phosphate across the cell membrane, and it has also been implicated in antibiotic resistance36. A single base pair deletion was observed in tRNA1Val (adenine37-N6)-methyltransferase in stavudine-resistant E. coli leading to truncation of 52% of the protein (Fig. 6). This particular methyltransferase methylates adenine in position 37 of tRNA1 (Val). Didanosine-resistant E. coli also exhibited four single base pair deletions in codon 50 of galactofuranose-binding protein (ytfQ), an ABC transporter involved in galactofuranose transport typically in carbon scavenging conditions but recently as a predictive gene for antimicrobial resistance37 (Fig. 6). The sodium/glutamate symporter possessed a single base pair insertion leading to the truncation of ten percent of the protein in dolutegravir-resistant E. coli. Although more work is needed to determine which of the genetic changes observed in the mutant genomes are important for the acquired antiviral or antimicrobial phenotypes, we have presented several likely candidates with a known role in antibiotic resistance, and which have led to the largest changes in protein sequences.
To investigate whether the mutations potentially implicated in antiviral resistance and antibiotic cross-resistance could be observed in a characterized multidrug-resistant E. coli model, we examined the publicly available full genome sequence for BAA-2471. When comparing multidrug-resistant E. coli BAA-2471 to antiviral-resistant E. coli strains, none of the mutations observed in antiviral-resistant E. coli strains leading to protein truncation were observed in BAA-2471 (Supplementary Data 14). For example, the mutations in thymidine kinase and bifunctional aspartate kinase/homoserine dehydrogenase that characterized zidovudine-resistant E. coli were not observed in BAA-2471. All the genes that contained insertions or deletions in antiviral-resistant E. coli mutants were intact for BAA-2471 and comparable to wild type E. coli.
Discussion
Despite the increasing public health concerns regarding the development and spread of antimicrobial resistance, the agents contributing to resistance development beyond antibiotic drugs remain incompletely understood21,38,39,40,41,42. Efforts to control antibiotic resistance are primarily focused on antibiotic stewardship for the prescription of antibiotic drugs43 but without attention to other drugs with antimicrobial properties. Antivirals have global widespread use, known antibacterial effects, and have even been considered as treatment for drug-resistant bacterial infections13,15. To date, no studies we are aware of have explored the potential for antiviral drugs to contribute to antimicrobial resistance. In this work, we examined the antibacterial activity of fourteen antiviral drugs of different antiviral classes to determine their potential to contribute to antiviral resistance and antibiotic cross-resistance. Our findings indicate that in addition to antibiotic drugs, antiviral drugs that exhibit antibacterial properties may also contribute to the antibiotic resistance burden.
Gram-negative E. coli and gram-positive B. cereus were chosen as well-characterized model organisms for investigation with environmental and clinical relevance44,45,46. Antivirals were tested at concentrations ranging from 0.1–100 μg/mL over 0–24 h (Fig. 1). The concentration range tested covers inhibitory and sub-inhibitory concentrations demonstrated in previous in vitro tests with bacteria10,11,12. We included concentrations tested in existing literature to ensure reproducible results and to validate our methods. The concentration range tested also includes therapeutic or circulating concentrations of antiviral drugs, such as the plasma concentration of zidovudine (0.016–1.7 mg/L)47 and therapeutic window of efavirenz (1–4 mg/L)48, as well as concentrations of antivirals detected in the aquatic environment that can reach concentrations in the μg/L range49. However, in attempt to establish inhibitory values for various incubation periods and exposure concentrations, the concentration ranges tested generally exceeded environmentally detected levels or a typical plasma concentration in the human body50.
We successfully isolated antiviral-resistant E. coli and B. cereus mutants after repeatedly exposing the bacteria to antiviral drugs with demonstrated antibacterial properties. When antiviral-resistant strains were challenged with antibiotic drugs and other antiviral drugs, each resistant strain exhibited a unique phenotypic profile of resistance to antiviral and antibiotic drugs. The resistance and cross-resistance profiles of the antiviral-resistant strains did seem to cluster by antiviral drug class, but none exhibited identical phenotypic resistance. The results suggest that despite structural similarity based on drug class in some cases, the antiviral drugs may be acting through different and possibly multiple pathways to exert their antibacterial properties. Resistant mutants therefore may possess a unique set of mutations conferring resistance to antiviral drugs and antibiotics.
In some cases, antiviral-resistance mutations conferred increased susceptibility to some antibiotics when resistant strains were challenged to test for cross-resistance. However, the increased sensitivity to particular classes of antibiotics in the dolutegravir- and raltegravir-resistant B. cereus strains, for example, may not be surprising when considered alongside the results of other studies that have observed increased susceptibility of certain drug-resistant strains of bacteria to other classes of antibiotic drugs28,51. Fitness costs and other changes in susceptibility due to resistance mutations have been described elsewhere as well39,52,53. The reduced maximal growth of zidovudine-resistant E. coli compared to wild type (Supplementary Data 4) and comparable growth to multidrug-resistant BAA-2471 (Fig. 5) may suggest how the mutations observed in this resistant strain come at a metabolic cost to the microorganism. However, the stability of the mutations throughout passaging even in the absence of drug suggest the mutation is favorable to sustain, or difficult to revert54 (Supplementary Data 7). Further investigation may help illuminate where and how the antiviral drugs exert their antimicrobial influence in cell replication or metabolism and how growth conditions also impact the effects of antimicrobials on bacteria55. The significance level of an...

Comments
Post a Comment