Intranasal immunization of mice with chimera of Salmonella Typhi protein elicits protective intestinal immunity | npj ... - Nature.com
Abstract
Development of safe, highly effective and affordable enteric fever vaccines is a global health priority. Live, oral typhoid vaccines induce strong mucosal immunity and long-term protection, but safety remains a concern. In contrast, efficacy wears off rapidly for injectable, polysaccharide-based vaccines, which elicit poor mucosal response. We previously reported Salmonella Typhi outer membrane protein, T2544 as a potential candidate for bivalent (S. Typhi and S. Paratyphi A) vaccine development. Here, we show that intranasal immunization with a subunit vaccine (chimera of T2544 and cholera toxin B subunit) induced strong systemic and intestinal mucosal immunity and protection from S. Typhi challenge in a mouse model. CTB-T2544 augmented gut-homing receptor expression on lymphocytes that produced Th1 and Th17 cytokines, secretory IgA in stool that inhibited bacterial motility and epithelial attachment, antibody recall response and affinity maturation with increased number of follicular helper T cells and CD4+ central and effector memory cells.
Similar content being viewed by others
Shigella virulence protein VirG is a broadly protective antigen and vaccine candidate
A bivalent vaccine confers immunogenicity and protection against Shigella flexneri and enterotoxigenic Escherichia coli infections in mice
Immunization of mice with chimeric antigens displaying selected epitopes confers protection against intestinal colonization and renal damage caused by Shiga toxin-producing Escherichia coli
Introduction
Salmonella enterica serovar Typhi (S. Typhi), a Gram-negative, intracellular pathogen and the etiological agent of typhoid fever, is a global threat to public health. The organism spreads through the faeco-oral route, causing significant morbidities and mortalities in the developing countries, especially of the south-east Asian region. As per Global Burden of Disease data, 9.24 million new typhoid fever cases occurred worldwide in 2019 (females 4.17 million and males 5.07 million), leading to 110,000 deaths and 8.05 million (3.86–13.9) disability adjusted life years (DALYs). Incidences in males and females were 7.3 (4.5–11.1) and 8.6 (5.4–12.9), respectively per 100,000 populations1. After crossing the intestinal barrier, Salmonella Typhi, being carried by the infected macrophages, spreads to the distant body sites like the liver, spleen, bone marrow and lymph nodes. As an obligate intracellular pathogen, the bacterium creates a niche for replication within the phagosomes of the infected macrophages, rendering it difficult for the host to eliminate it from the body2. The problem of S. Typhi infection is further complicated by the emergence and spread of multi-drug resistant and extensively drug-resistant (XDR) strains across continents3. While the mortality in untreated typhoid fever reaches nearly 20%, an incompletely cured acute infection may be followed by an asymptomatic chronic carrier state, which poses an increased risk for the development of adenocarcinoma of the gall bladder4.
Improvement of sanitation, potable water supply and food hygiene is considered essential for elimination of S. Typhi. However, vaccination may play a significant role in decreasing the incidence in the endemic countries in the short run. Two vaccines, namely a live, attenuated S. Typhi Ty21a strain for oral administration and an injectable preparation, containing purified Vi polysaccharide (Vi-PS) are widely available commercially. Despite high seroconversion rates with good short term protection, they are at best modestly efficacious in the long run5. Oral typhoid vaccine induces systemic and intestinal secretory antibodies as well as good T cell response, but requires at least three to four doses for optimum protection. Further, it is not recommended for children below 6 years of age due to difficulty in swallowing large capsules of the vaccine formulation. Live typhoid vaccine has also raised safety concerns because of occasional reports of bacteremia6. To escalate immunogenicity with a single dose of vaccine administration, several research groups have developed alternative attenuated strains in the S. Typhi Ty2 background. Several mutant vaccine strains, such as Ty800 with the deletion of the global regulator phoP/phoQ, M01ZH09/ZH09 lacking aroC and ssaV genes and double mutant (aroC and aroD) strain CVD908 have passed Phase II trials and showed good mucosal and serum antibody response in the volunteers. However, CVD 908 resulted in vaccinemia with a single oral dose of 107 viable organisms, and was later on modified by the deletion of another stress protein, htrA that prevented vaccinemia, but retained both humoral and cellular immune responses. To ensure more consistent serum anti-Vi antibodies, Vi was constitutively expressed in CVD 908 strain, generating CVD909. A prime boost regimen with orally administered CVD 909, followed by an injection of Vi-polysaccharide vaccine failed to induce strong Vi-specific antibody response, but instead, significantly raised Vi-specific IgA+ B memory cells. However, the need for pre-administration of buffer to neutralize stomach acid renders a potential delivery challenge for live attenuated vaccines5. None of the vaccine strains was shown to be effective against Paratyphoid infection.
Vi-polysaccharide vaccine, on the other hand, primarily works through the serum antibodies, but is poorly immunogenic with modest long-term efficacy, especially in children, because of T-cell-independent antibody response. Given the success of subunit vaccines for other bacterial infections, hunt for a highly efficacious typhoid vaccine, especially for smaller children (<2 years) that is safe and at the same time, capable of inducing durable protection is focused on developing typhoid conjugate (Vi-conjugate) vaccines (TCVs). A glycoconjugate vaccine (Vi-PS conjugated to tetanus toxoid, called Vi-TT), recently licensed in India has been reported to mount considerably high levels of durable protection through induction of memory B and T-cells7,8. The landmark TyVAC trial in Nepal, Malawi, and Bangladesh had shown protective efficacy of Typbar TCV to the extent of 79%, 84%, 85%, respectively9,10,11. However, protection conferred by Vi-TT and other TCVs, such as Vi-rEPA (polysaccharide conjugated to Pseudomonas aeruginosa exotoxin A), Vi-CRM197 and Vi-DT (diphtheria toxoid) would be exclusively mediated by the circulating anti-Vi antibodies due to the absence of intrinsic Salmonella proteins in these preparations12. Consequently, the emerging Vi-negative MDR (multidrug resistant) and XDR (extensive drug resistant) strains that were developed due to spontaneous genomic mutations in the regulatory regions of Vi polysaccharide of S. Typhi have evoked concerns12. There is a distinct threat that mass vaccination drive targeting Vi polysaccharide may exert a selection pressure for the existing Vi-negative S. Typhi and S. Paratyphi A and B strains, eventually rendering the vaccines to lose their protective efficacy. This has sparked an interest to generate future vaccines with universally present somatic antigens like T2544, which are immunogenic as well as essential for virulence of the bacteria. Antigen-specific, cytotoxic T cells, which were previously found to significantly augment anti-S. Typhi immunity through the elimination of intracellular bacteria were induced by recombinant T2544, as shown by our previous study, but not by the existing Vi conjugate vaccines13. Further, antibody secreting cells (ASCs), generated by Vi-PS and the TCVs will be primarily targeted to the systemic circulation, and not to the intestine14, whereas secretory antibodies, especially secretory IgA (sIgA) is believed to play a critical role in reducing systemic invasion of the luminal S. Typhi, thereby augmenting protection15.
Cholera toxin B subunit (CTB) has been extensively used as a vaccine adjuvant, especially for the protein antigens, owing to its nontoxic nature, ease of production of recombinant CTB and its fusion proteins in large quantities in E. coli and rapid renaturation of the recombinant proteins to the pentameric form of the native counterpart, enabling CTB to bind to its cognate GM1 ganglioside receptor16,17. This receptor is widely expressed by multiple, non-immune and immune cells (epithelial cells, macrophages, DCs and B cells), and CTB-binding to the receptor targets the antigens, co-administered or delivered as chimera to the desired cell populations, leading to augmented antigen uptake and presentation16,17. Other proposed mechanisms for the co-stimulatory effects of rCTB include direct lymphocyte activation18 and activation of the Toll-like receptors, although the identity of the TLRs remains unclear19. Adjuvanticity was most prominent when CTB was used as a genetically-fused partner of the vaccine antigen or chemically coupled to it, and delivered through the mucosal route. Indeed, CTB is one of the most potent mucosal adjuvants available to-date that attracted great attention20,21. Most antigens are poorly immunogenic when administered through the mucosal route, and tend to induce tolerance, especially at the intestinal mucosa. CTB often breaks the oral antigenic tolerance, but due to unknown reasons, significant oral adjuvanticity of CTB was only observed for live vaccines, such as oral cholera vaccine (Dukoral) that is already licensed for use22. In contrast, CTB administered through non-oral mucosal routes significantly enhanced antigen-specific humoral and cell-mediated immunity, not only at the local site, but also at distal mucosa, a phenomenon called "common mucosal immunity"23. Chemical and genetic conjugations of CTB with heterologous antigens from the simian immunodeficiency virus and Schistosoma mansoni have been proved to be potential vaccine candidates24,25. Adherent-invasive Escherichia coli (AIEC) tends to colonize the gut of patients with Crohn's disease and exacerbate inflammation. Intranasal immunization with AIEC siderophore enterobactin, conjugated to CTB induced robust mucosal antibody response against the siderophore in the small intestine and colon, thus reducing gut colonization of AIEC and colitis severity26. However, an earlier study showed that intestinal mucosal antibodies elicited against siderophores after intranasal CTB-siderophore conjugate administration could significantly reduce Salmonella colonization of the inflamed gut, but failed to reduce inflammation20.
We had previously reported that an outer membrane protein of S. Typhi Ty2 strain (T2544) is strongly immunogenic in mouse and naturally-infected humans27. A candidate vaccine formulation based on T2544 induced high levels of serum antibodies as well as robust effector and memory T cell responses, when administered into mice through the subcutaneous route, and protected them against S. Typhi challenge13,27. T2544 was present in multiple clinical isolates of S. Typhi and S. Paratyphi, while T2544-specific antibodies were detected in the acute and convalescent sera of patients infected with S. Typhi27.
Here, we report that intranasal immunization with purified CTB-T2544 was also protective against oral S. Typhi infection. The chimeric antigen augmented (i) secretory IgA in stool which inhibited bacterial motility and epithelial attachment (ii) affinity maturation of antibodies with strong recall response, (iii) gut-homing receptor expression on lymphocytes that produced Th1 and Th17 cytokines, and (iv) the number of follicular helper T cells and CD4+ effector memory cells. These results suggested that CTB-T2544 is a promising candidate for the development of an intranasal vaccine against human S. Typhi infection.
Results
Cloning, expression and purification of recombinant CTB-T2544 chimeric protein
To generate a gene chimera of ctx-b and t2544, 309 bp open reading frame (ORF) of V.cholerae ctx-b (cholera toxin B subunit) gene was amplified by PCR (Fig. S1a). In addition, ORF of Salmonella Typhi t2544 gene, along with the coding sequence of glycine-proline-glycine-proline (GPGP) linker at its NH2-terminus (total size 663 bp) was PCR-amplified (Fig. S1b). The amplicon of the ctx-b gene was cloned into the prokaryotic expression plasmid pET28a, using BamHI and SalI restriction sites, followed by cloning of the 663 bp amplicon of linker-t2544 at SalI and XhoI restriction sites immediately downstream of ctx-b (Fig. 1a). Confirmation of the fusion gene clone of ctx-b and t2544 was done by colony PCR (Fig. S1c), restriction digestion (Fig. 1b) and finally nucleotide sequencing (Fig. S1d). To generate the recombinant chimeric protein, pET28a plasmid construct carrying the ctx-b-t2544 fusion gene was transformed into the expression host E. coli BL21 (DE3) and the expression of the recombinant chimeric protein (rCTB-T2544) was induced by IPTG. rCTB-T2544 was purified by affinity chromatography using Ni-NTA agarose and the purity and specificity of the chimeric protein was confirmed by SDS-PAGE and western blots, respectively (Fig. 1c, d). Secondary structure of rCTB-T2544 derived from the analysis of the Far-UV circular dichroism (CD) spectra, using K2D2 (http://cbdm-01.zdv.uni-mainz.de/~andrade/k2d2/) indicated α-helical structure (72.38%) (Fig. S1e). Secondary structure of the other two proteins (rT2544 and rCTB) indicated 64.22% and 79.93% of α-helical structures, respectively. The yields of the purified rCTB-T2544, rCTB and rT2544 were 18.75 mg, 30 mg and 24 mg per liter of bacterial cultures, respectively. The endotoxin contaminations were found to be 0.015, 0.011, 0.008 EU/ml for rCTB, rT2544, and rCTB-T2544, respectively. Recombinant CTB-T2544, denatured by boiling, migrated in SDS-PAGE with the molecular weight marker that corresponded to the size of a chimera of T2544 and monomeric CTB (Fig. 1c; all the uncropped and unprocessed gel images are provided in Fig. S9). However, the role of CTB as an adjuvant requires a pentamer that is capable to bind to its cognate cell surface receptor, GM1 ganglioside. The size of the non-denatured rCTB-T2544, resolved in SDS-PAGE corresponded to a higher molecular weight protein containing pentameric CTB (Fig. 1e). To check if the native chimera indeed contained a CTB pentamer, we studied by ELISA the binding of rCTB-T2544 to the GM1 ganglioside receptor. The results showed that both the recombinant CTB and CTB-T2544 were able to bind to the receptor in a dose-dependent manner, suggesting the existence of CTB pentamer (Fig. 1f).
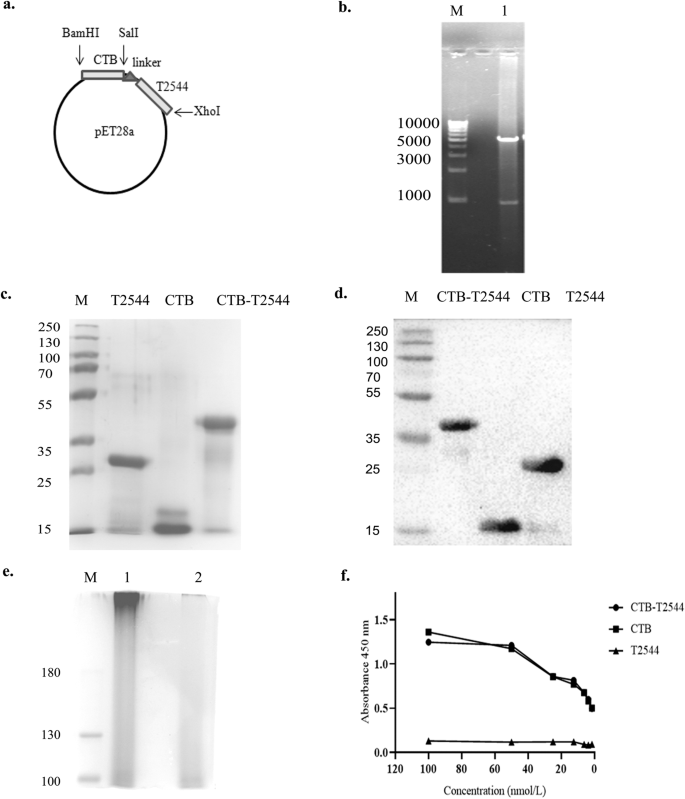
a Schematic diagram of cloned ctx-b and t2544 genes in pET28a vector. b 1% Agarose gel electrophoresis of pET28a-ctb-t2544 after restriction digestion with BamHI and XhoI in Lane 1; Lane-M shows 1 Kb plus DNA ladder. c Recombinant purified proteins run in 10% SDS–PAGE, followed by Coomassie blue staining; Lane-M, pre-stained protein marker. d Western blot analysis of purified recombinant proteins; the blot was developed using anti-HIS monoclonal antibody; Lane-M, pre-stained protein marker. e Denatured (boiled) and non-denatured (un-boiled) rCTB-T2544 protein, resolved in 10% SDS PAGE, followed by Coomassie blue staining of the gel. Lane M, Pre-stained protein marker, Lane 1, unboiled rCTB-T2544; Lane 2, rCTB-T2544 after boiling. All the blots or gels were derived from the same experiment and were processed in parallel. f GM1-ELISA after coating the plates with GM1 ganglioside (1 μg/well), followed by the addition of purified proteins as indicated. Plates were developed using anti-HIS antiserum (1:2500).
Immunization with rCTB-T2544 through the intranasal route protects against oral S. Typhi and Vibrio cholerae infections
The existence of the so-called "Common mucosal immune system"23 allows immunization through any mucosal route to induce immune response at the distant mucosal sites, albeit to a variable degree, due to migration of the activated immune cells. Intranasal immunization was earlier reported to elicit secretory IgA (sIgA) at the genital mucosa and salivary secretions24,25,26. To investigate if intranasal immunization with rCTB-T2544 induced protective immunity in the intestine, BALB/c mice were immunized with three doses of rCTB-T2544, rT2544, rCTB or the vehicle (PBS) at 12-day intervals (Fig. 2a). Twelve days after the 3rd dose, mice were challenged with 5 × 10^7 CFU (10 × LD50) of S. Typhi Ty2 strain through oral gavage (as mentioned under Methods section). An iron-overload mouse model of oral S. Typhi infection, established previously in our laboratory was used for the challenge experiment27. The results showed death of all the mice in the vehicle (PBS)-, rCTB- and rT2544- immunized groups within a period of 8 days, while 70% of mice that received CTB-T2544 were alive after 30 days of infection (Fig. 2b). Further, when the immunized animals were subjected to an ileal loop experiment with the administration of cholera toxin28, fluid accumulation in the loops was significantly less, resulting in lower loop weight/length ratio for rCTB- and rCTB-T2544-immunized animals compared with the ones that received T2544 or the vehicle (Fig. 2c, d). Together, the above results underscored the efficacy of rCTB-T2544 as a candidate bivalent vaccine for S. Typhi and V. cholerae infections.
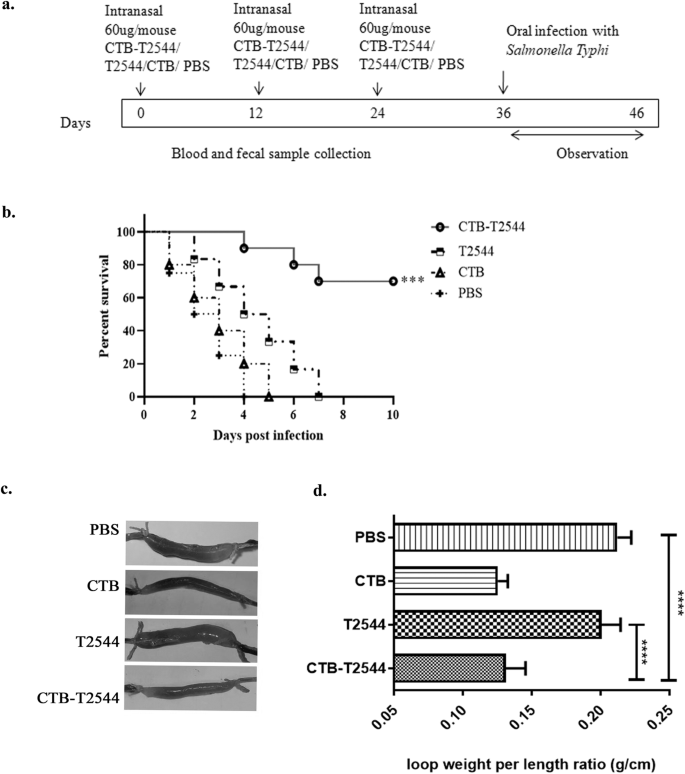
a Immunization schedule, sample collection, and bacterial challenge of BALB/c mice. Blood and stool samples were collected before each immunization dose and 12 days after the last immunization. b Mouse survival assay. Kaplan-Meyer plot of cumulative mortality of mice immunized intranasally with the vehicle or the recombinant protein antigens. Mice (n = 10/group) were challenged with S. Typhi 12 days after the last immunization dose and were monitored for 10 days. Significance was calculated by comparing the survival of the mice immunized by rCTB-T2544 with T2544 by Log-Rank Mantel Cox test. ***p = 0.0001 c In parallel experiments, mice immunized as above were subjected to ileal loop assay by injecting 100 ng of cholera toxin into each loop. The animals (n = 8/group) were checked for fluid accumulation in the ileal loops after 8 hours. d Fluid accumulation was measured by calculating the weight divided by the length of individual loops. Significance was calculated by comparing the loop weight/ length ratio of CTB-T2544 group with the T2544 and PBS immunized group, using one-way ANOVA with post hoc analysis using Tukey's multiple comparison test. Data represented as mean with SD; ****p value < 0.0001 vs. T2544 and PBS. Error bars represent SD.
Intranasal immunization with rCTB-T2544 induces protective serum and mucosal antibody response
To check if CTB could augment humoral immune response to the co-administered antigen T2544, antibody end-point titers (The reciprocal of the titer, at which the absorbance of the sample was the same as those from PBS-immunized mice) were determined by ELISA 12 days after completion of primary immunization dose. Markedly raised titers of T2544-specific serum IgG, IgG1, IgG2a, and IgA antibodies were observed in the mice immunized with rCTB-T2544, as compared with the rT2544-immunized mice (Fig. 3a). Similarly, CTB-specific IgG and IgA antibody titers were also raised in the serum (Fig. S3a). These results were in agreement with the recovery of substantial numbers of T2544-specific IgG and IgA antibody-secreting cells (ASCs) in the spleen, MLN, and Peyer's Patches (PP) after intranasal rCTB-T2544, as analyzed by the enzyme-linked immunosorbent spot (ELISpot) assays, while ASCs were much fewer in the other groups of mice (Figs. 3b, S3b).

a ELISA plates were coated with rT2544 protein and serum T2544-specific antibody titers were determined. Samples were serially half-diluted, from 1:200 to 1:25600 for IgG and 1:20 to 1:2560 for IgA. Optical density at 450 nm was recorded. The reciprocal of the titer, at which the absorbance of the sample was the same as the unimmunized mice samples was plotted in log2 scale. b Two weeks after the primary immunization, mice were sacrificed and cells were isolated from the spleen, MLN and Peyer's Patches (PP). ELISPOT assay was performed with anti-mouse IgG, IgA and total immunoglobulin. c T2544-specific IgA antibodies in the fecal extracts and intestinal contents were quantified two weeks after the primary immunization. d Adhesion Inhibition Assay. Bacteria were preincubated with 1:50 dilution of the serum, fecal extracts or intestinal wash samples of the unimmunized and immunized mice for 30 min and used for adhesion assay as described under Methods. Mean CFU recovered from the lysed cells were plotted. e Opsonophagoctosis assay. Bacteria were pre-incubated with 1:50 dilution of the serum, fecal contents or intestinal lavage, followed by infection of the THP-1 cell monolayer. Opsonophagocytosis assay was performed as described under Methods. Phagocytosed bacteria were calculated after cell lysis and mean fold increase for the immunized samples compared with the pre-immune counterparts was plotted. f Motility inhibition assay. Motility of Salmonella Typhi Ty2 in LB agar (0.4%), containing 5% serum, fecal extract or intestinal contents from the immunized mice was measured by the zone diameter of bacterial growth after 6 h. The above experiments were repeated three times and mean of the values from all three experiments were plotted. Error bars represent SD. A representative picture from the motility assays is shown. Significance was calculated using two tailed unpaired t test, comparing CTB-T2544 group with the corresponding T2544 group. **** p < 0.0001, ***p = 0.0002.
Intestinal sIgA antibody could significantly contribute to the protection against S. Typhi, an enteric pathogen. We measured antigen-specific sIgA titers in the stool and intestinal contents and found strong T2544-specific sIgA response only in the mice immunized with rCTB-T2544 (Fig. 3c), which also mounted CTB-specific sIgA (Fig. S3c). Protective efficacies of the serum and secretory antibodies were revealed by the inhibition of S. Typhi adherence to the intestinal epithelial cell monolayers, impairment of bacterial motility in the soft agar and the promotion of opsanophagocytosis. Sera, fecal extracts and intestinal contents from the experimental mice were pre-incubated with S. Typhi, which was subsequently used to infect HT-29 and THP-1 cell monolayers. Adhesion of the bacteria to HT-29 cell monolayer was studied by CFU counts and confocal microscopy. Bacterial adhesion to the cells was inhibited by the antibodies present in different samples, out of which maximum inhibition was found for the samples collected from rCTB-T2544 immunized mice (Figs. 3d, S3d). We also observed that a 1:5000 dilution of the rCTB-T2544 antibodies had comparable adhesion to 1:50 dilution of the pre-immunization samples, indicating that the former had 100-fold higher functional competence for adhesion inhibition (Fig. S6). Similarly, in the opsanophagocytosis assay, mean peak fold increase of bacterial phagocytosis by THP-1 cells was significantly higher following opsonization of the bacteria with antibodies from rCTB-T2544-immunized mice compared with the other groups of animals (Figs. 3e, S3e). Finally, we evaluated soft agar motility inhibition of S. Typhi after incubation with the above antibodies and found much greater inhibition when antibodies from the mice immunized with the chimeric protein were used (Fig. 3f). Together, the above results suggested that intranasal immunization with a CTB chimera of S. Typhi antigen (T2544) induced greater number of ASCs specific to the antigen in the secondary lymphoid tissues and higher levels of antigen-specific circulating and mucosal antibodies with greater protective efficacies against S. Typhi infection, when compared with the antigen administered without CTB. To further dissect relative contribution of the serum and mucosal antibodies, we performed adoptive transfer of pooled immune sera from rCTB-T2544-immunized mice to naïve mice, followed by a lethal bacterial challenge. This resulted in a 25% survival rate, suggesting a relatively minor role played by the serum antibodies in overall protection (Fig. S5a). To study the contribution from mucosal antibodies in the protection, we pre-incubated S. Typhi with intestinal lavage and fecal extracts from the immunized mice for 30 minutes before infecting the naïve mice. A lethal dose killed only 50% mice, while with a sublethal dose of the pre-incubated bacteria showed significant reduction in the colonization of the intestine (ileum, cecum, colon Fig. S5 b–d) and spread to the internal visceral organs (MLN, spleen and liver Fig. S5e–g) compared with the similar dose of bacteria that were not pre-incubated with mucosal antibodies. These findings highlighted a more significant role of mucosal antibodies in imparting protection.
Intranasal immunization with rCTB-T2544 increased the recruitment of DCs and lymphocytes expressing gut-homing receptors to the MLN
Lung DCs were previously reported to imprint gut homing receptors on naïve (IgM+) B cells and CD4+ T cells in the local (mediastinal) lymph nodes after intranasal antigen administration, in addition to class switch recombination of B cells, generating IgG+ and IgA+ cells29. However, other studies had suggested that tissue homing of naïve lymphocytes is coupled with their site of activation by the DCs, and only the lymphocytes primed in the MLNs express α4β7 receptor that is required for migration to the intestine, although splenic lymphocytes show promiscuity with regard to tissue homing30. To address this issue in our study, we checked for DC recruitment to the MLNs of the immunized mice. The percentage of CD11c+ DCs in the MLNs after rCTB-T2544 immunization was nearly four times higher than after immunization with rT2544 (26.7% vs 7.0%) (Fig. 4a), indicating that intranasal CTB promotes DC migration to the draining nodes of the intestine. Next, to address DC migration, migratory DC (mDC) markers (MHChiCD11CintCD103+) expression in the MLN DCs was assessed and found to be 3-fold higher (Fig. S7) in the MLN of the rCTB-T2544 immunized mice compared with the vehicle-immunized mice. Increased number of MLN DCs could result in greater number of lymphocytes expressing gut homing receptors. Indeed, we found significantly higher number of B and T cells, not only in the MLNs, but also from the spleen expressing α4β7 and CCR9 receptors (Figs. 4b, c, S4). DCs play a critical role in mucosal sIgA production by expressing several factors like BAFF, APRIL, iNOS, RALDH1, TGF-β and IL-631. Given increased DC migration to the MLNs and strong sIgA response in the intestine after intranasal immunization with CTB-T2544, we studied for the IgA-inducing factor expression by the MLN DCs. The results showed augmented expression of the factors as above by the MLN DCs (Fig. 4d). Together the above results suggested that intranasal CTB upregulates the expression of IgA-inducing factors on the local DCs and promotes their migration to the MLNs. These MLN DCs imprint gut homing receptors on the lymphocytes, which then migrate to the intestinal mucosa to execute the effector function by sIgA and cytokines production.
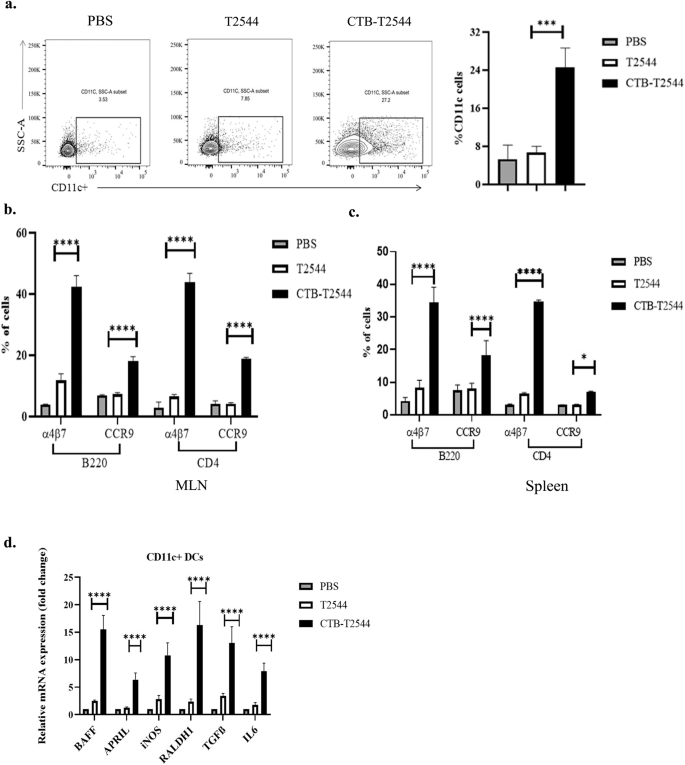
a Twelve days after the primary immunization, MLN cells were surface-stained with CD11c and analyzed by flow cytometry. b, c Gut homing receptor expression on B and T lymphocytes. Twelve days after the primary immunization, lymphocytes isolated from the MLN (b) and spleen (c) of the experimental mice were stained with fluorochrome-tagged antibodies against cell lineage markers (CD4 and B220) and gut homing receptors (α4β7 and CCR9) and analyzed by flow cytometry. Data presented as percentages of cells out of the total T and B cells, expressing the homing receptors. d RT-qPCR analysis of gene expressions in DCs, isolated from the MLN of experimental mice were plotted. GAPDH-normalized expression of all the genes was represented as fold changes of the PBS-immunized group. All experiments were repeated three times, and data from a representative one are presented in (a) and mean ( ± SD) of the data from all three experiments are shown under (b–d). Significance was calculated using one way ANOVA (a) and two-way ANOVA (b,c) with post hoc analysis using Tukey's multiple comparison test. Error bar represents SD. *p = 0.0281, ***p = 0.0008 and ****p < 0.0001 (T2544 vs. CTB-T2544).
CTB-T2544 immunization induces a balanced T helper cell response
Th1 response is required for the clearance of intracellular pathogens like Salmonella32, while Th2 and Th17 cells promote serum antibodies and sIgA production, respectively. We found significantly elevated, circulating Th1 (IL12 and IFNγ) and Th2 (IL-4, IL-5, IL-10) cytokines in rCTB-T2544-immunized mice as opposed to only modest elevation in the comparator immunized groups (Fig. 5a). In addition, the number of IFNγ- and IL-17A-secreting CD4+ T cells (Th1 and Th17 cells, respectively) in the Peyer's Patches was also increased markedly after CTB-T2544 immunization (Fig. 5b). We also checked if CTB induced follicular helper T (TFH) cells, which play a crucial role in the germinal center formation and the development of high-affinity antibodies and memory B cells33. TFH cell number in MLN after immunization with rCTB-T2544 was doubled compared with rT2544 or no immunization (6.03% vs 3.06% vs 2.74%, respectively) (Fig. 5c). Together, these results suggested that rCTB-T2544 chimera helped to create a protective milieu in the intestine through the induction of T cell subsets and cytokines.
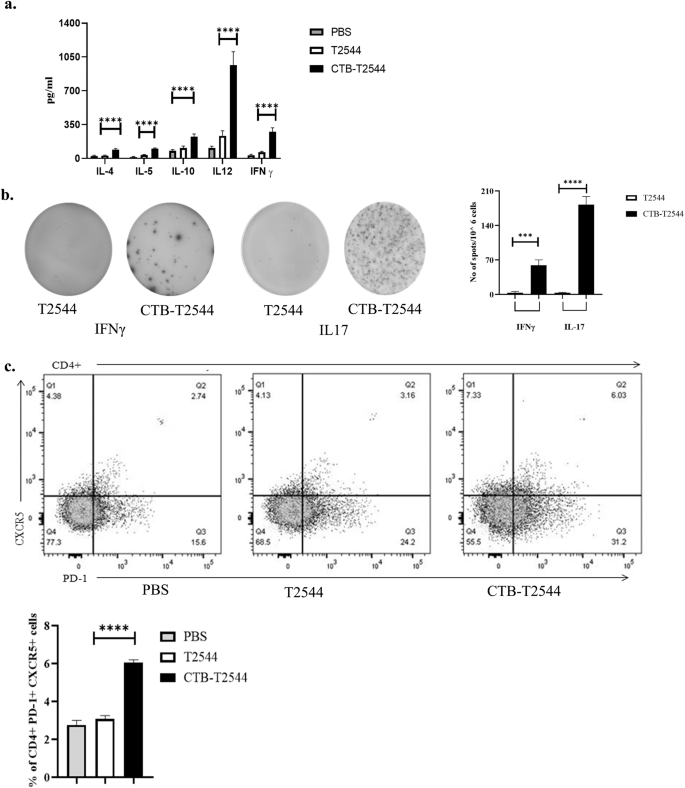
a Th1 (IL-12, IFN-γ), Th2 (IL-4, IL-5), and Treg (IL-10) cell cytokines in the serum samples of the unimmunized and immunized mice were measured by ELISA. b Cells from the PP of the experimental mice were incubated with 10 µg of T2544 for 24 h. Cytokine-secreting cells were enumerated by ELISpot assays and data presented as spot forming cells/106 total cells analyzed. c Analysis of TFH (T follicular helper) cells in the MLN. Cells were detected by measuring the expression of CXCR5 and PD-1 on CD4+ cells using flow cytometer. Significance was calculated by one way (b, c) and two-way (a) ANOVA with post hoc analysis using Tukey's multiple comparison test. The experiment was repeated three times and representative images from one experiment are shown. Bar diagram represents the statistical data from all three experiments. Error bars represent SD. ****p < 0.0001, ***p = 0.0006.
CTB-T2544 immunization elicits memory response
Antigen-specific memory T cell response was evaluated by co-culture of the CD4+ T-cells, isolated from the spleen of the experimental mice with S. Typhi-pulsed bone marrow dendritic cells (BMDCs) derived from naïve mice. IFNγ release measured in the culture supernatants was significantly higher for the CD4+ cells recovered from the mice immunized with rCTB-T2544 as compared with the animals that received rT2544 or left unimmunized, suggesting augmentation of the memory response by CTB (Fig. 6a). With respect to specific memory cell subsets, percentages of both central memory (CD62LhighCD44high) and effector memory (CD62LlowCD44high) T cells (TCM and TEM)34 were increased after immunization with the chimeric antigen containing CTB (Fig. 6b). In contrast, these mice had half the number of naïve CD4+ cells in the spleen as compared with the other immunized groups. To compare the induction of memory B cells after immunization, a booster dose was administered on 108th day of experiment and serum anti-T2544 antibodies were measured 12 days later. A steeper rise of antibody production with significantly higher titers was observed after rCTB-T2544 immunization as compared with the immunization using rT2544 alone (Fig. 6c), suggesting the critical role played by CTB in antigen-induced differentiation of memory B cells into the plasma cells, producing IgG. Avidity index of anti-T2544 IgG antibodies was assessed by measuring the OD of the ELISA plates after washing the immune complexes with or without 4 M urea-containing buffer. We observed significantly high avidity index (60–65%) after the primary immunization, which further went up to 70–78% following the booster immunization (Fig. 6d), suggesting successful priming. In contrast, intranasal T2544 immunization resulted in poor priming (avidity index < 35%). These results together suggest that intranasal rCTB-T2544 may elicit long-term immunity against S. Typhi infection.

a Memory T cells were analyzed by ELISA for IFN-γ release in the culture supernatants of CD4+ T cells, isolated from the spleen of the experimental mice and co-cultured with BMDCs, pulsed with rT2544. Data represents mean( ± SD) of three independent experiments. b The percentages of TEM (CD4+CD44hiCD62Llo) and TCM (CD4+CD44hiCD62Lhi) cells from the spleens of different experimental groups were quantified using flow cytometry. Representative images from one of the experiments and statistical analysis of the data from all three experiments are shown. c Serum antibody titers (mean ± SD) measured in the immunized mice (n = 6) at the indicated time points are shown. Arrows indicate primary (3 doses at 0, 12 and 24th day) and booster (108th day) immunization. Mean ± SD were plotted for CTB-T2544 and T2544. d Avidity assay. Anti-T2544 avidity index was measured by ELISA and represented as the ratio of IgG bound to T2544 in the presence and absence of 4 M urea, multiplied by 100. Significance was calculated using one way (a, d) and two-way (b) ANOVA with post hoc analysis using Tukey's multiple comparison test. Error bars represent SD. ****p < 0.0001.

Comments
Post a Comment